
Биология. 11 класс
§ 23. Генетический код и его свойства
Как вы знаете, признаки и свойства каждого организма определяются прежде всего белками, которые синтезируются в его клетках. Белки выполняют самые разнообразные функции (вспомните какие), обеспечивая тем самым протекание процессов жизнедеятельности. Можно сказать, что именно от этих биополимеров в первую очередь и зависит существование организма. Однако время функционирования белков, как и многих других биомолекул, весьма ограничено. Поэтому синтез белков в организме должен осуществляться непрерывно. Этот процесс протекает во всех клетках одноклеточных и многоклеточных организмов.
Вам также известно, что хранителем наследственной (генетической) информации, т. е. информации о первичной структуре белков, является ДНК. Участок молекулы ДНК, содержащий информацию о первичной структуре одного белка, получил название ген. Кроме того, генами называют участки ДНК, хранящие информацию о строении молекул рРНК и тРНК.
В биосинтезе белков, который осуществляется в рибосомах, ДНК прямого участия не принимает. Передача генетической информации, содержащейся в ДНК, к месту синтеза белка происходит с помощью посредника. Этим посредником является матричная (информационная) РНК (мРНК, иРНК), которая синтезируется на одной из цепей молекулы ДНК по принципу комплементарности.
В молекулах ДНК и мРНК информация о первичной структуре белков «записана» в виде последовательности нуклеотидов. Сами же белки синтезируются из аминокислот. Значит, в природе существует особая система кодирования, на основании которой последовательность нуклеотидов расшифровывается в виде последовательности аминокислот молекул белков. Этот «шифр» называется генетическим кодом. Таким образом, генетический код — это система записи информации о первичной структуре белков в виде последовательности нуклеотидов ДНК (мРНК).
Генетический код обладает следующими свойствами.
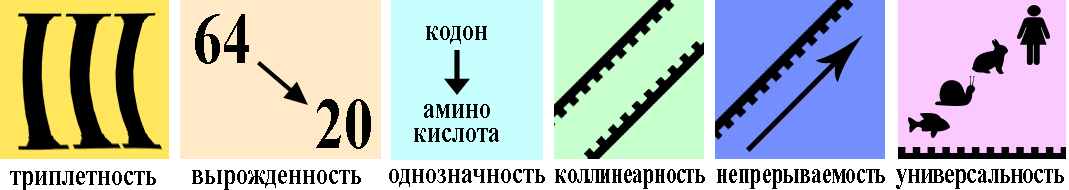
1. Код является триплетным. Это значит, что каждая аминокислота кодируется триплетом (кодоном) — сочетанием трех последовательно расположенных нуклеотидов. В состав молекул ДНК и РНК входит по 4 типа нуклеотидов. Если бы за определенную аминокислоту «отвечал» один нуклеотид, можно было бы закодировать только 4 из 20 белокобразующих аминокислот. Дублетов (по два нуклеотида) хватило бы лишь на 4 2 = 16 аминокислот. Количество возможных триплетов (сочетаний трех нуклеотидов) составляет 4 3 = 64. Этого с избытком хватает для кодирования всех 20 видов аминокислот (табл. 23.1).
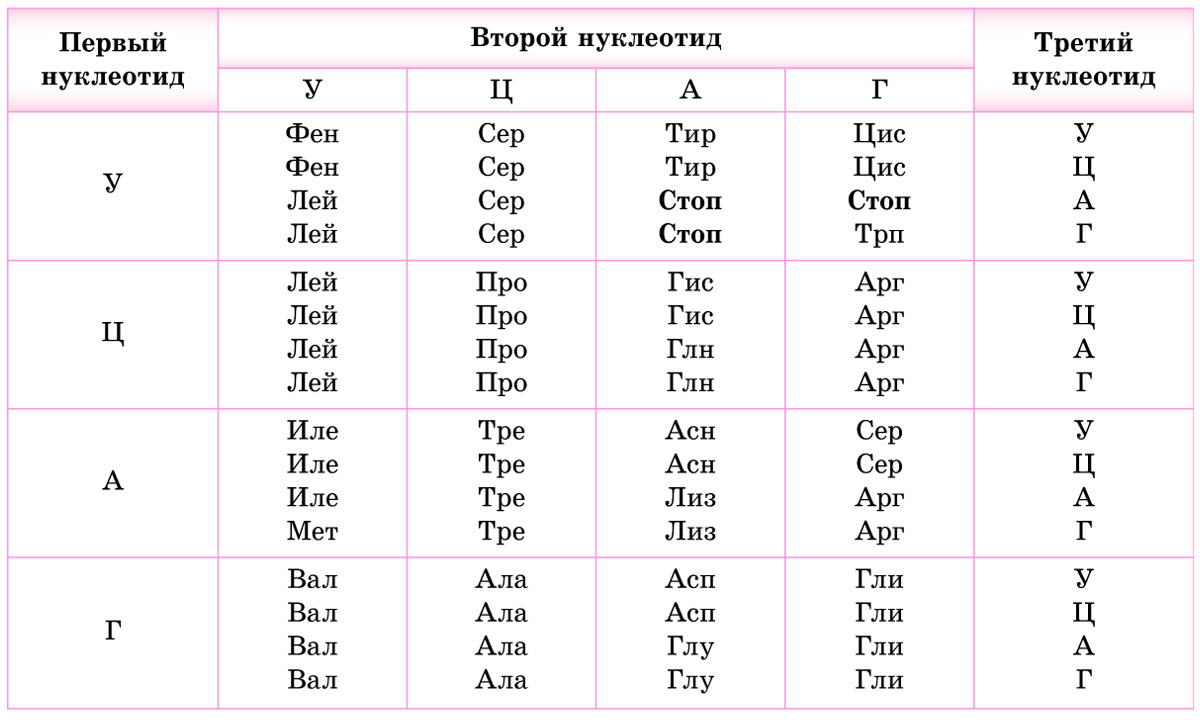
Обратите внимание, что 3 из 64 кодонов (в молекулах мРНК — УАА, УАГ и УГА) не кодируют аминокислоты. Это так называемые стоп-кодоны *или нонсенс-кодоны (от англ. nonsense — бессмыслица)*, они служат сигналом окончания синтеза белка. *Остальные триплеты называются смысловыми.*
* Генетический код расшифровали американские биохимики Р. Холли, Х. Г. Корана и М. Ниренберг в середине прошлого века. Работа стартовала в 1961 г. В бесклеточные системы, содержащие все необходимые компоненты для синтеза белка (рибосомы, аминокислоты, тРНК и др.), ученые сначала вводили искусственно синтезированные мРНК, состоящие только из одного типа нуклеотидов. Было выяснено, что в присутствии, например, полицитидиловой мРНК (ЦЦЦЦЦЦ. ) синтезируется полипептид, состоящий только из остатков аминокислоты пролина, в присутствии полиуридиловой (УУУУУУ. ) — из фенилаланина. Стало понятно, что кодону ЦЦЦ соответствует пролин, а триплет УУУ кодирует фенилаланин. К 1965 г., благодаря использованию искусственно синтезированных молекул мРНК с известными повторяющимися последовательностями нуклеотидов, удалось расшифровать все остальные триплеты. В 1968 г. это открытие было удостоено Нобелевской премии.*
2. Код однозначен — каждый триплет кодирует только одну аминокислоту.
3. Как уже отмечалось, число триплетов превышает количество кодируемых аминокислот. Поэтому генетический код является избыточным (вырожденным) — одна и та же аминокислота может кодироваться разными триплетами. Например, в мРНК цистеин (Цис) может быть закодирован триплетом УГУ или УГЦ, треонин (Тре) — АЦУ, АЦЦ, АЦА или АЦГ. Некоторые аминокислоты, например лейцин (Лей), кодируются шестью различными триплетами, в то же время метионину (Мет) и триптофану (Трп) соответствует только по одному кодону (проверьте по таблице генетического кода).
4. Код не перекрывается — один и тот же нуклеотид не может одновременно входить в состав двух соседних триплетов.
5. Код непрерывен. В полинуклеотидной цепи нуклеотиды располагаются непрерывно и соседние триплеты ничем не отделены друг от друга. Это значит, что фактически деление на триплеты условно — все зависит от того, с какого именно нуклеотида начинается их считывание. Поэтому в клетках считывание информации, содержащейся в генах, всегда начинается со строго определенного нуклеотида.
Если в составе гена происходит изменение количества нуклеотидов (их выпадение или вставка) на число, не кратное трем, наблюдается так называемый сдвиг рамки считывания (рис. 23.1). Это прив одит к существенному изменению последовательности аминокислот в белке, который кодируется измененным геном. В некоторых случаях сдвиг рамки считывания приводит к возникновению стоп-кодонов, из-за чего синтез белка обрывается.
*Суть происходящего при сдвиге рамки считывания можно понять на следующем примере. Прочитайте предложение, составленное из трехбуквенных слов (аналогично триплетам):
ЖИЛ БЫЛ КОТ ТИХ БЫЛ СЕР МИЛ МНЕ ТОТ КОТ.
В этом предложении заключен определенный смысл, понять который можно и без знаков препинания. Выпадение одной буквы аналогично выпадению одного нуклеотида. Оно приводит к изменению порядка считывания и потере смысла:
ЖЛБ ЫЛК ОТТ ИХБ ЫЛС ЕРМ ИЛМ НЕТ ОТК ОТ — выпадение второй буквы.
То же самое произошло бы и после вставки лишней буквы. В случае замены одной буквы либо при изменении их количества на три смысл предложения меняется не столь значительно. Например:
ЖИВ БЫЛ КОТ ТИХ БЫЛ СЕР МИЛ МНЕ ТОТ КОТ — замена третьей буквы;
БЫЛ КОТ ТИХ БЫЛ СЕР МИЛ МНЕ ТОТ КОТ — выпадение первых трех букв.
Однако смысл предложения (в нашей аналогии — первичная структура белка) во многом зависит от положения измененных букв (нуклеотидов). Так, смысл может существенно исказиться:
ЖИЛ БОТ ТИХ БЫЛ СЕР МИЛ МНЕ ТОТ КОТ — выпадение пятой, шестой и седьмой букв.
Аналогичная ситуация наблюдается и с белками. В зависимости от расположения замененной (утраченной, добавленной) аминокислоты молекула белка может сохранить пространственную конфигурацию и функции, частично изменить их или же полностью утратить свои исходные характеристики.*
Как уже отмечалось, правильное считывание генетической информации обеспечивается только тогда, когда оно начинается со строго определенной позиции. У эукариот стартовым кодоном молекулы мРНК является триплет АУГ. Именно с него и начинается считывание.
6. Код универсален — у всех живых организмов одним и тем же триплетам соответствуют одни и те же аминокислоты. Иными словами, у всех организмов генетический код расшифровывается одинаково (за редким исключением). Это свидетельствует о единстве происхождения живых организмов.
*Некоторые вариации генетического кода обнаружены у бактерий, инфузорий, дрожжей, в коде митохондриальной ДНК и т. д. Например, у бактерий триплет мРНК ГУГ может играть роль стартового кодона, а у эукариот он предназначен только для кодирования аминокислоты валин. В митохондриях млекопитающих триплет УГА кодирует триптофан, в то время как в матричной РНК, синтезированной в ядре клетки, он служит стоп-кодоном. И наоборот, в коде митохондрий триплеты АГА и АГГ являются сигналами окончания синтеза белка, а в «основной версии» генетического кода им соответствует аминокислота аргинин.*
Биосинтез белка и нуклеиновых кислот. Гены, генетический код
В обмене веществ организма ведущая роль принадлежит белкам и нуклеиновым кислотам.
Белковые вещества составляют основу всех жизненно важных структур клетки, обладают необычайно высокой реакционной способностью, наделены каталитическими функциями.
Нуклеиновые кислоты входят в состав важнейшего органа клетки — ядра, а также цитоплазмы, рибосом, митохондрий и т. д. Нуклеиновые кислоты играют важную, первостепенную роль в наследственности, изменчивости организма, в синтезе белка.
План синтеза белка хранится в ядре клетки, а непосредственно синтез происходит вне ядра, поэтому необходима служба доставки закодированного плана из ядра к месту синтеза. Такую службу доставки исполняют молекулы РНК.
После дальнейших изменений этот вид закодированной и-РНК готов. и-РНК выходит из ядра и направляется к месту синтеза белка, где буквы и-РНК расшифровываются. Каждый набор из трех букв и-РНК образует «букву», обозначающее одну конкретную аминокислоту.
Другой вид РНК отыскивает эту аминокислоту, захватывает ее с помощью фермента и доставляет к месту синтеза белка. Эта РНК называется транспортной, или т-РНК. По мере прочтения и перевода сообщения и-РНК цепочка аминокислот растет. Эта цепочка закручивается и укладывается в уникальную форму, создавая один вид белка. Примечателен даже процесс укладки белка: на то, чтобы с помощью компьютера просчитать все варианты укладки белка среднего размера, состоящего из 100 аминокислот, потребовалось бы 1027 (!) лет. А для образования в организме цепочки из 20 аминокислот требуется не более одной секунды, и этот процесс происходит непрерывно во всех клетках тела.
Гены, генетический код и его свойства.
На Земле живет около 7 млрд людей. Если не считать 25—30 млн пар однояйцовых близнецов, то генетически все люди разные : каждый уникален, обладает неповторимыми наследственными особенностями, свойствами характера, способностями, темпераментом.
Такие различия объясняются различиями в генотипах—наборах генов организма; у каждого он уникален. Генетические признаки конкретного организма воплощаются в белках — следовательно, и строение белка одного человека отличается, хотя и совсем немного, от белка другого человека.
Ген – единица наследственной информации организма, которой соответствует отдельный участок ДНК
Генетический код состоит из троек (триплетов) нуклеотидов ДНК, комбинирующихся в разной последовательности (ААТ, ГЦА, АЦГ, ТГЦ и т.д.), каждый из которых кодирует определенную аминокислоту (которая будет встроена в полипептидную цепь).
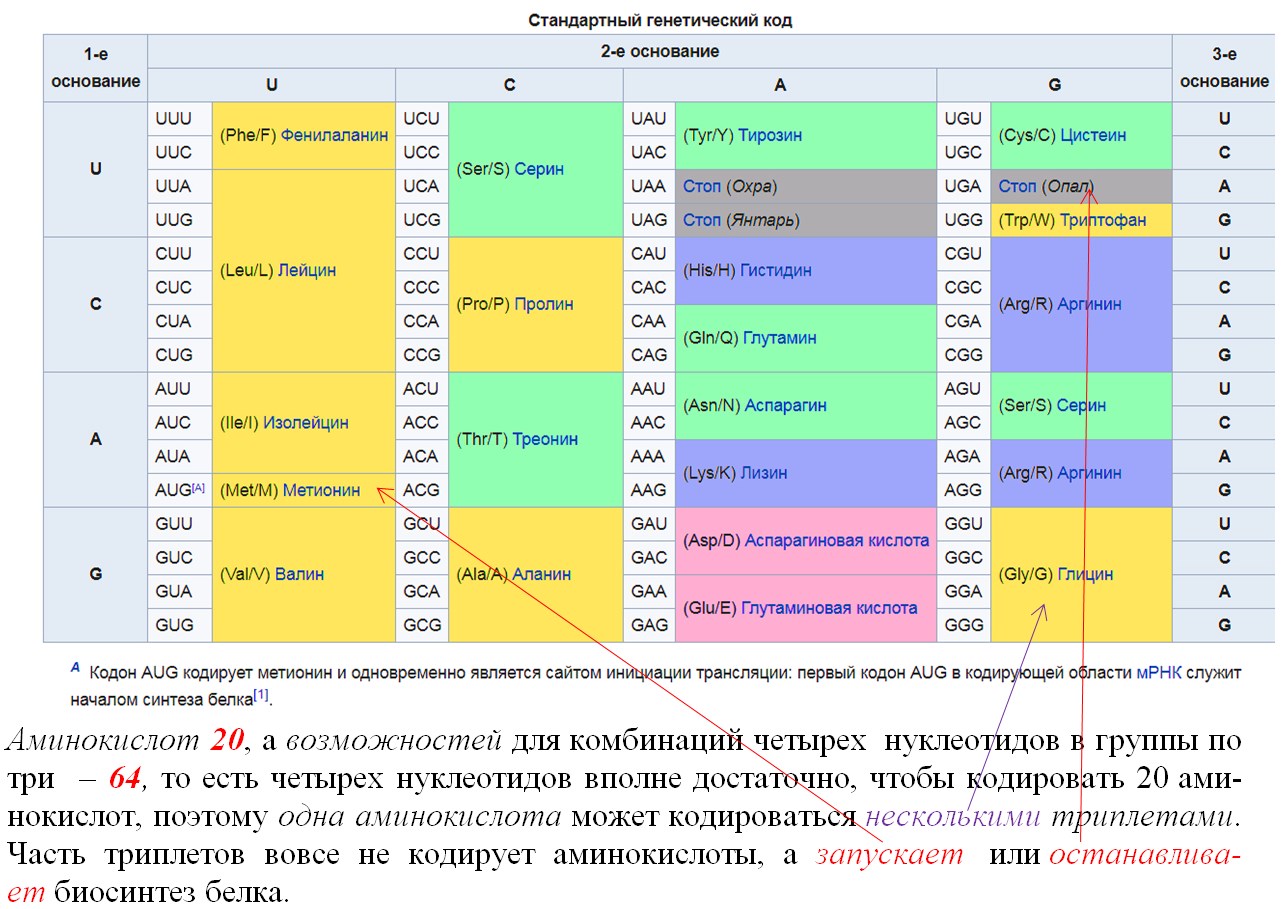
Основные свойства генетического кода:
2. Избыточность ( вырожденность ) кода является следствием его триплетности и означает то, что одна аминокислота может кодироваться несколькими триплетами (поскольку аминокислот 20, а триплетов — 64), за исключением метионина и триптофана, которые кодируются только одним триплетом. Кроме того, некоторые триплеты выполняют специфические функции: в молекуле и-РНК триплеты УАА, УАГ, УГА — являются терминирующими кодонами, т. е. стоп-сигналами, прекращающими синтез полипептидной цепи. Триплет, соответствующий метионину (АУГ), стоящий в начале цепи ДНК, не кодирует аминокислоту, а выполняет функцию инициирования (возбуждения) считывания.
4. Коллинеарность кода, т.е. последовательность нуклеотидов в гене точно соответствует последовательности аминокислот в белке.
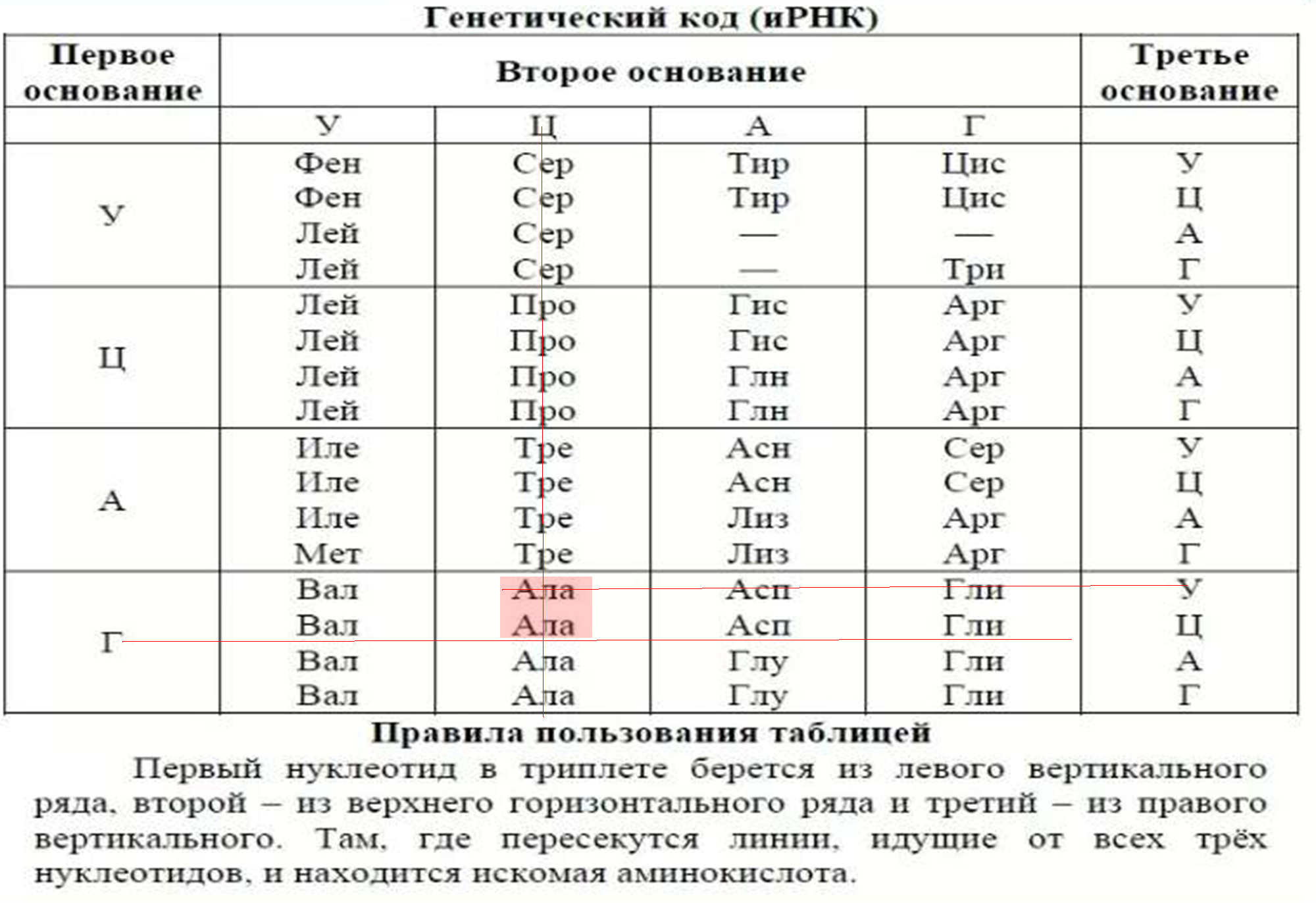
Существуют таблицы генетического кода для расшифровки кодонов и- РНК и построения цепочек белковых молекул.
Реакции матричного синтеза.
В живых системах встречается реакции, неизвестные в неживой природе — реакции матричного синтеза.
Термином «матрица» в технике обозначают форму, употребляемую для отливки монет, медалей, типографского шрифта: затвердевший металл в точности воспроизводит все детали формы, служившей для отливки. Матричный синтез напоминает отливку на матрице: новые молекулы синтезируются в точном соответствии с планом, заложенным в структуре уже существующих молекул.
Матричный принцип лежит в основе важнейших синтетических реакций клетки, та-ких, как синтез нуклеиновых кислот и белков. В этих реакциях обеспечивается точная, строго специфичная последовательность мономерных звеньев в синтезируемых полимерах.
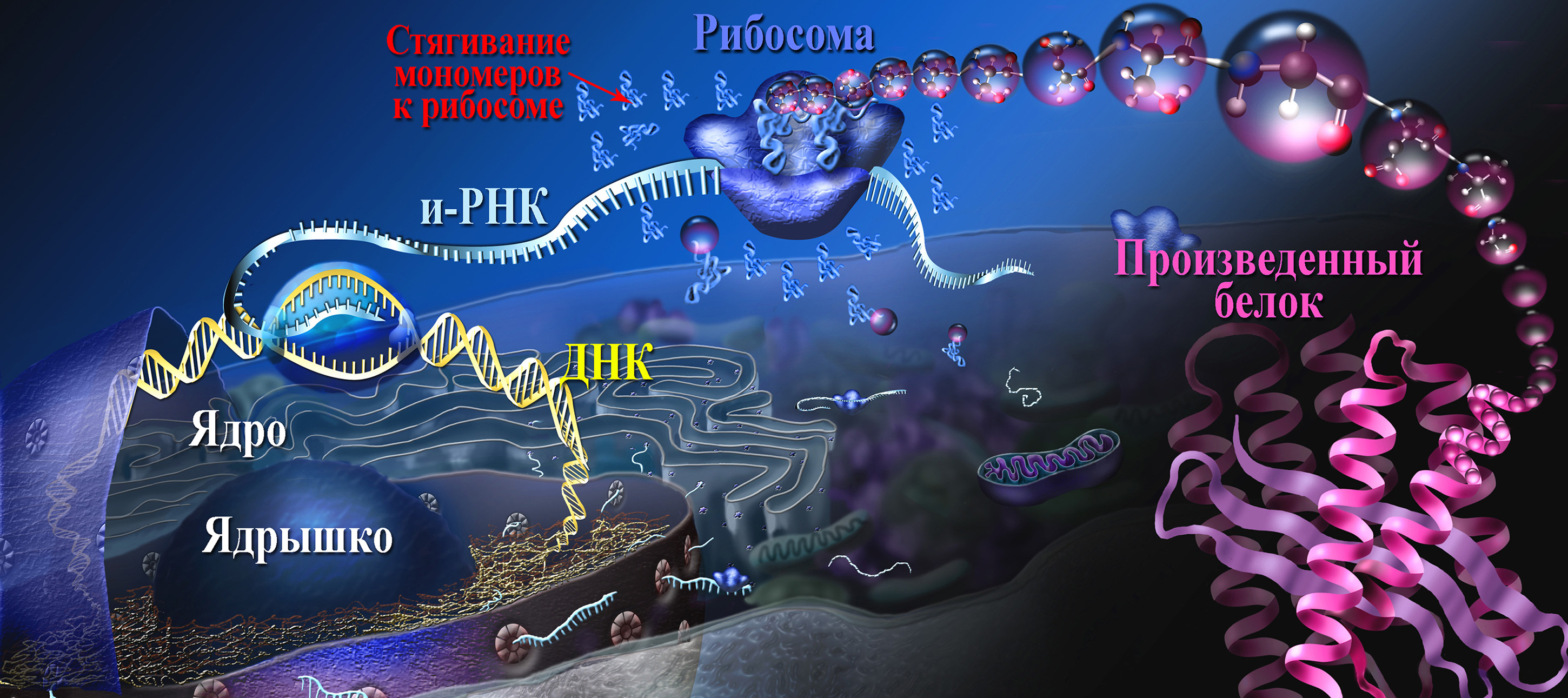
Мономерные молекулы, из которых синтезируется полимер, — нуклеотиды или аминокислоты — в соответствии с принципом комплементарности располагаются и фиксируются на матрице в строго определенном, заданном порядке.
Затем происходит «сшивание» мономерных звеньев в полимерную цепь, и готовый полимер сбрасывается с матрицы.
После этого матрица готова к сборке новой полимерной молекулы. Понятно, что как на данной форме может производиться отливка только какой-то одной монеты, одной буквы, так и на данной матричной молекуле может идти «сборка» только какого-то одного полимера.
Матричный тип реакций — специфическая особенность химизма живых систем. Они являются основой фундаментального свойства всего живого — его способности к воспроизведению себе подобного.
Реакции матричного синтеза
Молекула ДНК состоит из двух комплементарных цепей. Эти цепи удерживаются слабыми водородными связями, способными разрываться под действием ферментов. Молекула ДНК способна к самоудвоению (репликации), причем на каждой старой половине молекулы синтезируется новая ее половина.
Кроме того, на молекуле ДНК может синтезироваться молекула и-РНК, которая затем переносит полученную от ДНК информацию к месту синтеза белка.
Передача информации и синтез белка идут по матричному принципу, сравнимому с работой печатного станка в типографии. Информация от ДНК многократно копируется. Если при копировании произойдут ошибки, то они повторятся во всех последующих копиях.
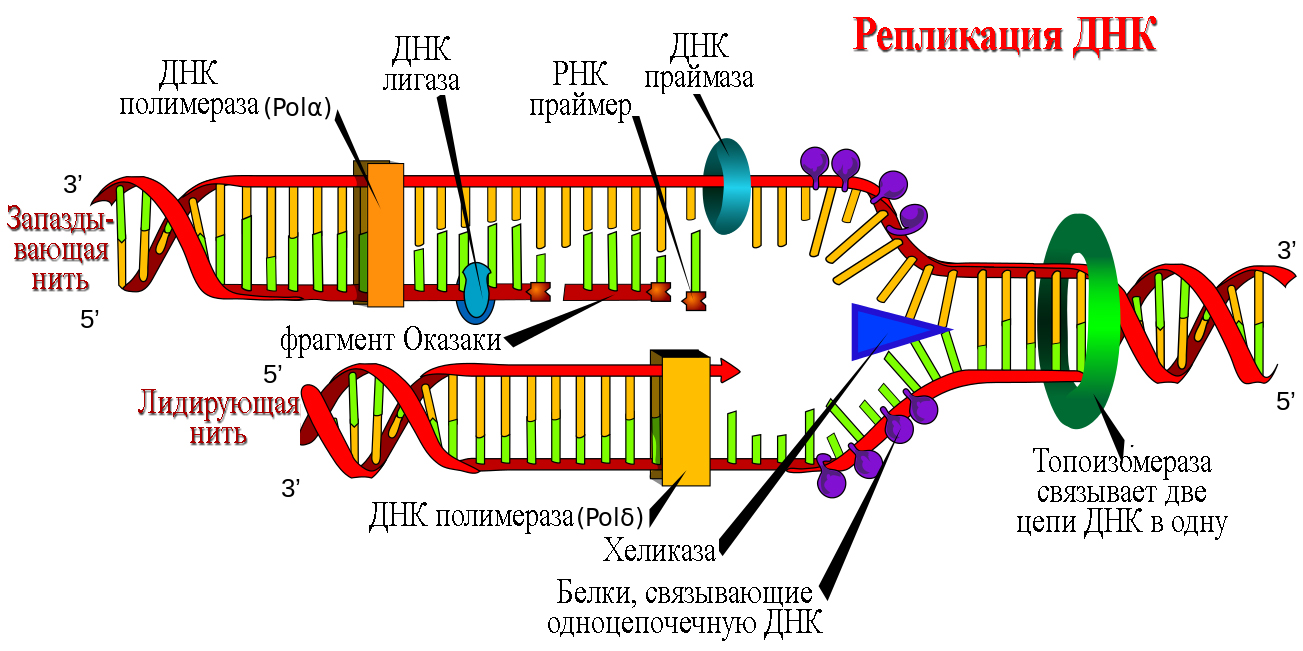
Правда, некоторые ошибки при копировании информации молекулой ДНК могут исправляться — процесс устранения ошибок называется репарацией. Первой из реакций в процессе передачи информации является репликация молекулы ДНК и синтез новых цепей ДНК.
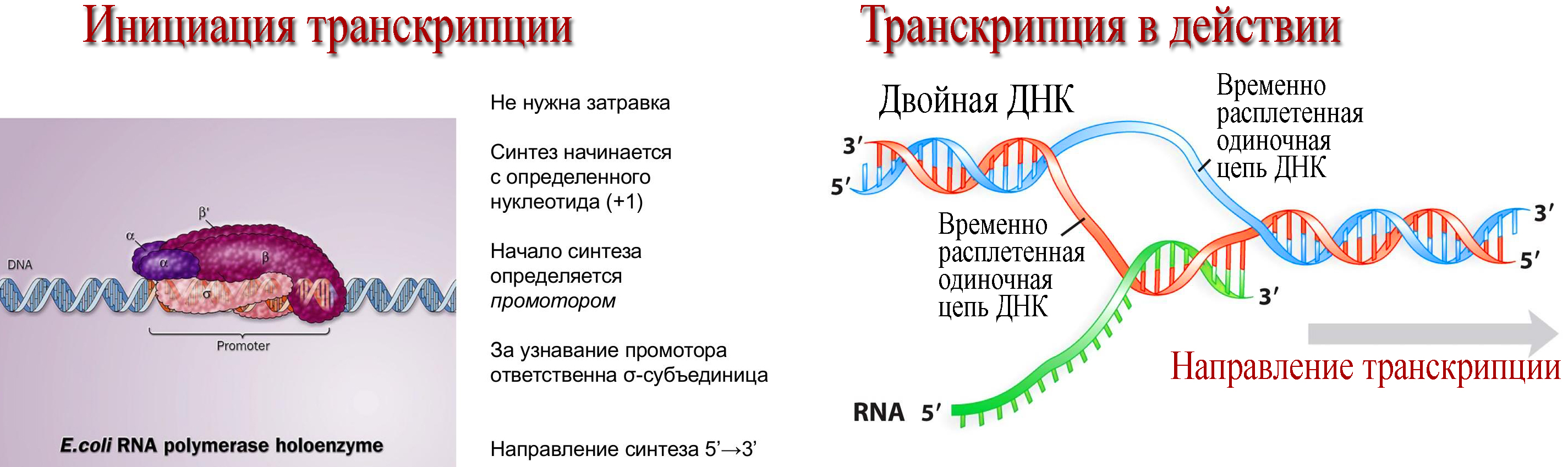
2. Транскрипция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Готовая молекула и-РНК выходит в цитоплазму на рибосомы, где происходит синтез полипептидных цепей.
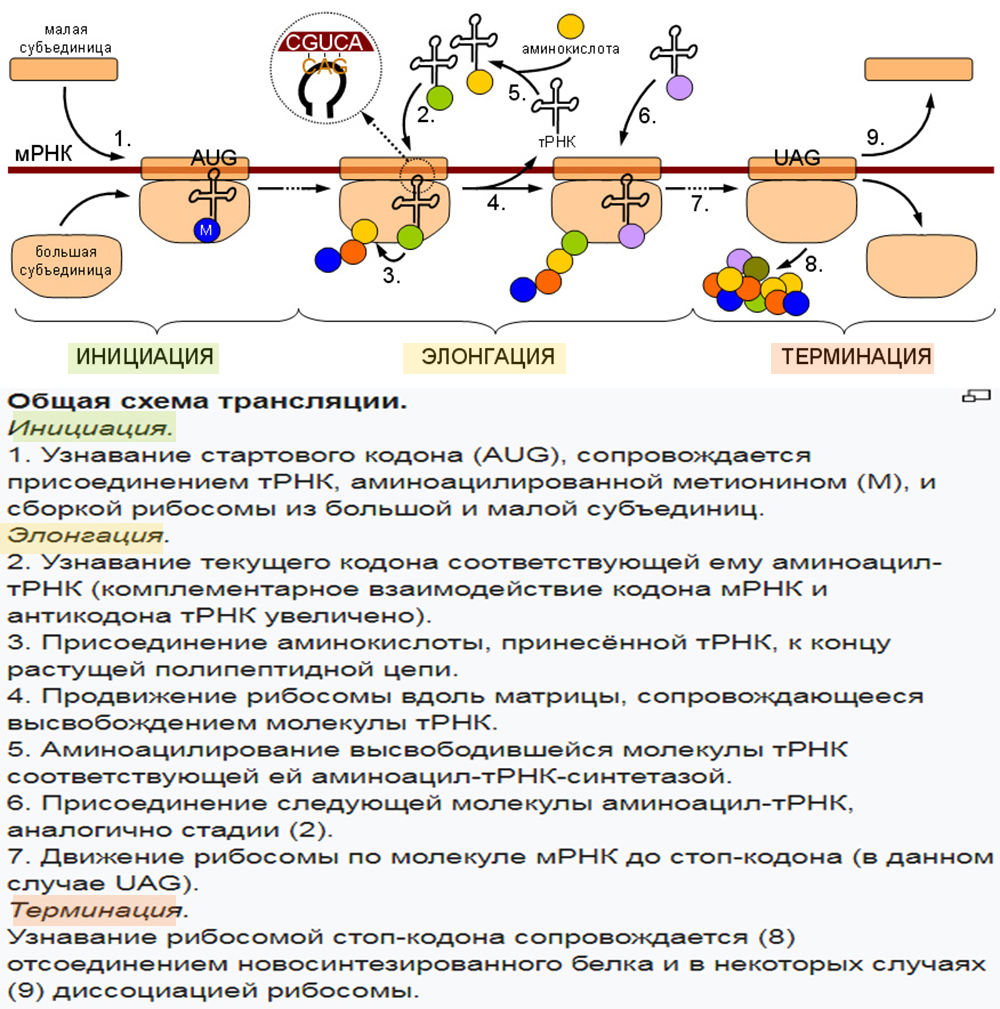
3. Трансляция (от лат. translatio — перенос, перемещение) — процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК), осуществляемый рибосомой. Иными словами, это процесс перевода информации, со-держащейся в последовательности нуклеотидов и-РНК, в последовательность амино-кислот в полипептиде.
4. Обратная транскрипция — это процесс образования двуцепочечной ДНК на основании информации из одноцепочечной РНК. Данный процесс называется обратной транскрипцией, так как передача генетической информации при этом происходит в «обратном», относительно транскрипции, направлении. Идея обратной транскрипции вначале была очень непопулярна, так как противоречила центральной догме молекулярной биологии, которая предполагала, что ДНК транскрибируется в РНК и далее транслируется в белки.
Последовательность матричных реакций при биосинтезе белков можно представить в виде схемы.
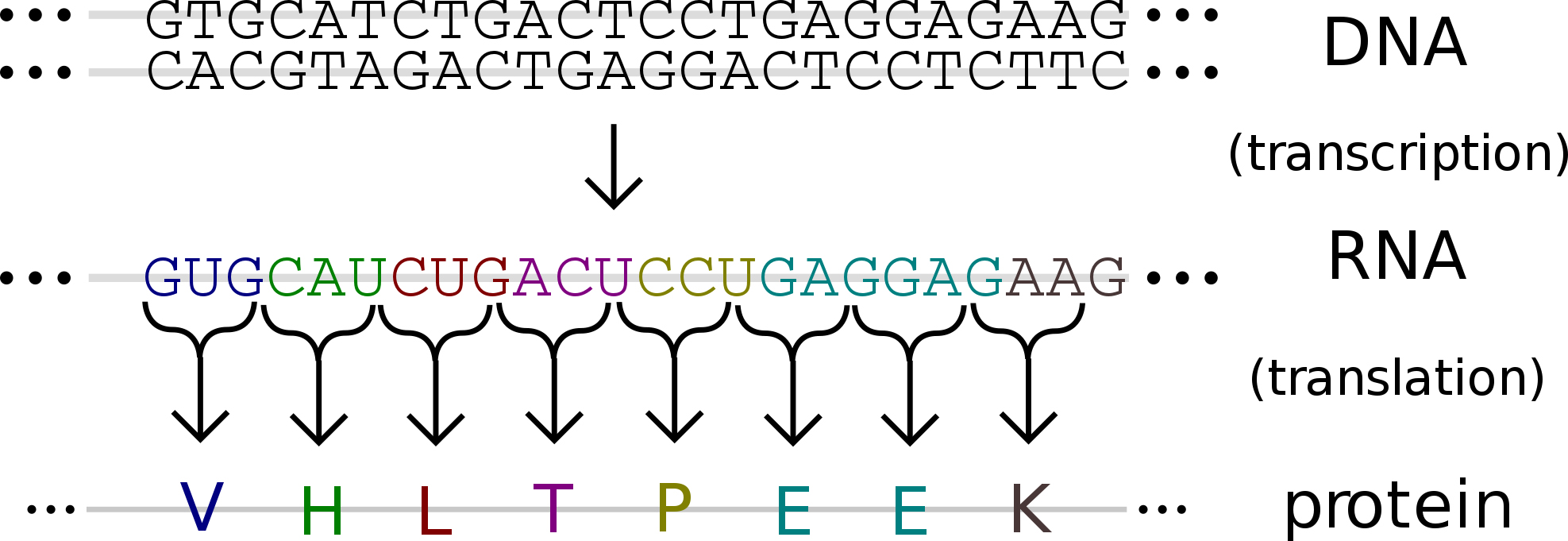
Таким образом, биосинтез белка – это один из видов пластического обмена, в ходе которого наследственная информация, закодированная в генах ДНК, реализуется в определенную последовательность аминокислот в белковых молекулах.
В результате активирования аминокислота становится более лабильной и под действием того же фермента связывается с т-РНК. Каждой аминокислоте соответствует строго специфическая т-РНК, которая находит «свою» аминокислоту и переносит ее в рибосому.
Следовательно, в рибосому поступают различные активированные аминокислоты, соединенные со своими т-РНК. Рибосома представляет собой как бы конвейер для сборки цепочки белка из поступающих в него различных аминокислот.
Одновременно с т-РНК, на которой «сидит» своя аминокислота, в рибосому поступает «сигнал» от ДНК, которая содержится в ядре. В соответствии с этим сигналом в рибосоме синтезируется тот или иной белок.
Направляющее влияние ДНК на синтез белка осуществляется не непосредственно, а с помощью особого посредника – матричной или информационной РНК (м-РНК или и-РНК), которая синтезируется в ядре под влиянием ДНК, поэтому ее состав отражает состав ДНК. Молекула РНК представляет собой как бы слепок с формы ДНК. Синтезированная и-РНК поступает в рибосому и как бы передает этой структуре план — в каком порядке должны соединяться друг с другом поступившие в рибосому активированные аминокислоты, чтобы синтезировался определенный белок. Иначе, генетическая информация, закодированная в ДНК, передается на и- РНК и далее на белок.
Молекула и-РНК поступает в рибосому и прошивает ее. Тот ее отрезок, который находится в данный момент в рибосоме, определенный кодоном (триплет), взаимо-действует совершенно специфично с подходящим к нему по строению триплетом (антикодоном) в транспортной РНК, которая принесла в рибосому аминокислоту.
Транспортная РНК со своей аминокислотой подходит к определенному кодону и-РНК и соединяется с ним; к следующему, соседнему участку и- РНК присоединяется другая т-РНК с другой аминокислотой и так до тех пор, пока не будет считана вся цепочка и-РНК, пока не нанижутся все аминокислоты в соответствующем порядке, образуя молекулу белка. А т-РНК, которая доставила аминокислоту к определенному участку полипептидной цепи, освобождается от своей аминокислоты и выходит из рибосомы.
Затем снова в цитоплазме к ней может присоединиться нужная аминокислота, и она снова перенесет ее в рибосому. В процессе синтеза белка участвует одновременно не одна, а несколько рибосом — полирибосомы.
Основные этапы передачи генетической информации:
Этапы универсальны для всех живых существ, но временные и пространственные взаимоотношения этих процессов различаются у про- и эукариотов.
У прокариот транскрипция и трансляция могут осуществляться одновременно, поскольку ДНК находится в цитоплазме. У эукариот транскрипция и трансляция строго разделены в пространстве и времени: синтез различных РНК происходит в ядре, после чего молекулы РНК должны покинуть пределы ядра, пройдя через ядерную мембрану. Затем в цитоплазме РНК транспортируются к месту синтеза белка.
Генетический код допускает разночтения

Генетический код основан на однозначном соответствии каждого кодона (тройки нуклеотидов) определенной аминокислоте. Это правило, казавшееся нерушимым, не выполняется у инфузории Euplotes, у которой в пределах одного и того же гена кодон UGA может кодировать две разных аминокислоты — цистеин или селеноцистеин. Подлинный смысл каждого конкретного кодона UGA определяется специальной регуляторной последовательностью нуклеотидов в некодирующей части гена.
Селеноцистеин (selenocysteine) — нестандартная «двадцать первая» аминокислота, отличающаяся от обычного цистеина тем, что вместо атома серы в ее состав входит атом селена. Селеноцистеин является обязательным компонентом нескольких важных ферментов в организме животных (включая человека), простейших, бактерий и архей. Белки, содержащие селеноцистеин, называют селенопротеинами (selenoproteins). В геноме человека содержится более 20 генов селенопротеинов, поэтому селен является необходимым компонентом питания, и его недостаток в пище приводит к различным заболеваниям.
Стандартный генетический код, однако, предусматривает возможность кодирования только двадцати «канонических» аминокислот, и для того, чтобы закодировать двадцать первую, эволюции пришлось пойти на хитрость. Для кодирования селеноцистеина используется кодон UGA, который по умолчанию является стоп-кодоном, то есть сигнализирует об окончании синтеза белковой молекулы. Однако если за кодирующим участком гена следует особая последовательность нуклеотидов, которая называется SECIS (selenocysteine insertion sequence), аппарат синтеза белка интерпретирует кодон UGA как кодирующий селеноцистеин.
У организмов, в геноме которых есть гены селенопротеинов, имеется специальная селенопротеиновая транспортная РНК, которая распознает кодон UGA при помощи комплементарного ему антикодона UCA. Последовательность SECIS в ходе транскрипции включается в состав матричной РНК, синтезируемой на основе селенопротеинового гена. Оказавшись в составе матричной РНК, SECIS сворачивается в особую трехмерную структуру — «шпильку» с двумя петельками (см. рисунок), которая и служит сигналом, заставляющим аппарат синтеза белка присоединить к кодону UGA транспортную РНК, несущую селеноцистеин.
Антон Туранов и Вадим Гладышев из Университета штата Небраска (Линкольн, США) и их коллеги из нескольких американских научных институтов изучили механизм кодирования селеноцистеина у инфузории Euplotes. Эта инфузория интересна тем, что у нее, в отличие от других инфузорий и подавляющего большинства живых существ, кодон UGA не является стоп-кодоном и «по умолчанию» кодирует цистеин. Исследователи подумали, что если у Euplotes есть селенопротеины, то эта инфузория может оказаться единственным организмом, у которого один и тот же триплет нуклеотидов кодирует, в зависимости от контекста, две разные аминокислоты.


Так и оказалось. Исследователи обнаружили в геноме Euplotes восемь генов селенопротеинов, в каждом из которых имеется последовательность SECIS и как минимум один кодон UGA. В четырех из этих генов обнаружилось более одного такого кодона. Как выяснилось, только один из UGA-кодонов в каждом гене кодирует селеноцистеин. Остальные интерпретируются клеткой как кодирующие цистеин. Таким образом, кодон UGA у Euplotes может кодировать две разные аминокислоты даже в пределах одного и того же гена.

В ядерном геноме Euplotes найдено целых три гена транспортных РНК, распознающих кодон UGA: селеноцистеиновая тРНК и два варианта цистеиновой тРНК. В митохондриальном геноме Euplotes кодон UGA кодирует триптофан, и в соответствии с этим имеется еще четвертая, митохондриальная триптофановая тРНК, распознающая этот кодон.
Чтобы проверить, насколько универсальным является механизм кодирования селеноцистеина у разных организмов, исследователи пересадили селенопротеиновые гены инфузории (вместе с последовательностями SECIS) в человеческие эмбриональные клетки. Оказалось, что человеческий аппарат синтеза белка правильно понимает смысл тех кодонов UGA в генах инфузории, которые кодируют селеноцистеин. Человеческие клетки успешно синтезировали селенопротеины на основе генов инфузории, используя при этом человеческую селеноцистеиновую тРНК. Однако это произошло только с теми селенопротеиновыми генами инфузории, в которых кодон UGA один, и он кодирует селеноцистеин. Наткнувшись на кодон UGA, кодирующий у инфузории цистеин, человеческие клетки интерпретировали его как стоп-кодон и прекращали синтез белковой молекулы. Что и понятно, ведь у человека нет цистеиновых тРНК, распознающих кодон UGA.
Исследователи также попытались выяснить, каким образом клетка понимает, какие из кодонов UGA в пределах данного гена следует интерпретировать как цистеиновые, а какие — как селеноцистеиновые. Как выяснилось, это зависит от структуры SECIS. Каждый вариант последовательности SECIS указывает клетке на строго определенный участок гена длиной в несколько десятков нуклеотидов, в пределах которого все кодоны UGA интерпретируются как селеноцистеиновые. За пределами этого участка кодоны UGA трактуются в соответствии с их значением «по умолчанию»: у инфузорий — как цистеиновые, у человека — как стоп-кодоны.
От структуры последовательности SECIS зависит, на каком расстоянии от конца гена будет располагаться участок, в пределах которого все UGA будут транслироваться в селеноцистеин. Значение тонких различий в структуре SECIS понимается одинаково инфузориями и человеческими клетками. Чтобы выяснить всё это, исследователи конструировали и вставляли в человеческие клетки различные модифицированные варианты селенопротеиновых генов (передвигали с места на место кодоны UGA, заменяли один вариант SECIS на другой и т. д.).
Таким образом, генетический код, «в норме» используемый для кодирования 20 аминокислот, при необходимости может быть расширен. Кроме селеноцистеина, который считают 21-й аминокислотой, существует и 22-я — пирролизин (pyrrolysine). Он входит в состав некоторых важных ферментов у организмов, считающихся одними из древнейших на планете, — архей-метаногенов (см. метаногенез). Пирролизин, как и селеноцистеин, кодируется каноническим стоп-кодоном, но другим (UAG), и тоже в сочетании со специальной регуляторной «шпилькой» в некодирующей области гена.
Может быть, эти странные неканонические способы кодирования аминокислот помогут пролить свет на тайну происхождения генетического кода. В статье об этом не говорится, но ведь можно предположить, что в ходе эволюции механизмов специфического синтеза полипептидов в РНК-мире строгое и однозначное соответствие между кодонами и аминокислотами сформировалось не сразу, и поначалу весь генетический код держался «на шпильках».
Источник: Anton A. Turanov, Alexey V. Lobanov, Dmitri E. Fomenko, Hilary G. Morrison, Mitchell L. Sogin, Lawrence A. Klobutcher, Dolph L. Hatfield, Vadim N. Gladyshev. Genetic Code Supports Targeted Insertion of Two Amino Acids by One Codon // Science. 2008. V. 323. P. 259–261.
При такой ненависти к науке даже странно (и немного досадно), что вы успели научиться пользоваться клавиатурой.
Редакция полностью поддерживает Александра Маркова. Мы хотим еще подчеркнуть, что это не форум, где можно приятно поболтать с понравивишимся тебе собеседником (как почему-то думают некоторые пользователи), а комментарии к конкретным новостям. Поэтому все записи, которые не имеют прямого отношения к данной новости, будут удаляться, а пользователи, злоупотребляющие такой болтовней, блокироваться.